
In den ersten beiden Folgen der Serie über generelle Merkmale der Fossilüberlieferung der Lebewesen wurden einige Vorhersagen zusammengestellt, die zutreffen sollten, falls es eine allgemeine Evolution der Lebewesen gegeben haben sollte. In dieser dritten und letzten Folge geht es darum, ob sich die folgenden beiden Erwartungen von Evolutionsbiologen in der Fossilüberlieferung auch bestätigen:
• Gibt es tatsächlich ungezählte Übergangsformen (bzw. Bindeglieder) zwischen Arten oder grösseren Gruppen?
• Und lassen sich dementsprechend die fossil bekannten als auch die heute existierenden Arten wirklich in Form eines eingeschachtelten Systems darstellen?
Laut Evolutionsmodell sollten sich alle bekannten heutigen und fossilen Arten widerspruchsfrei in ein solches System einordnen lassen. Wenn das gelingt, werten es Evolutionsbiologen als eine Art «Evolutionsbeweis» (siehe dazu die erste Folge dieser Serie).
Bei vielen Tier- und Pflanzengruppen ist ein solch eingeschachteltes System tatsächlich erkennbar, allerdings ist es in unterschiedlichem Ausmass «gestört». Das heisst: Viele Arten oder ganze Gruppen lassen sich bezüglich ihrer Merkmalskonstellationen und Ähnlichkeiten nicht in eine eingeschachtelte Form bringen. Wir werden im Folgenden einige Beispiele kennenlernen.

NOCH GAR NICHT ENTDECKT?
Für die Rekonstruktion eines eingeschachtelten Systems beziehungsweise eines evolutionären Stammbaums, benötigt man eine gewaltige Zahl an Übergangsformen oder Bindeglieder, die zwischen den bekannten Formen schrittweise vermitteln. Daher gehen Evolutionsbiologen davon aus, dass wir 99 bis 99,997 Prozent aller fossilen Arten, die es in diesem Stammbaum gegeben haben müsste, noch gar nicht entdeckt haben. 1 Allerdings können Formen nur dann als geeignete Bindeglieder gelten, wenn ihr gesamter Merkmalsmix widerspruchsfrei zwischen zwei andere Arten oder grössere Gruppen passt. Anders ausgedrückt: Ein optimales Bindeglied sollte Merkmale dieser beiden Arten oder Gruppen kombinieren, aber so gut wie keine sonstigen anderen komplexen Merkmale besitzen.
“Haben wir 99 bis 99,997 Prozent aller fossilen Arten noch gar nicht entdeckt?
Ob also bestimmte Zwischenformen (d. h. die neutrale Formulierung ohne Deutung) als evolutionäre Übergangsformen (bzw. Bindeglieder) interpretiert werden können, muss in jedem Fall eigens begründet werden. Mosaikformen hingegen, die Merkmale verschiedener Gruppen kombinieren, sind nicht automatisch als Übergangsformen interpretierbar. Ein lehrreiches Beispiel ist das Schnabeltier (Abb. 1).
Die Frage nach der Existenz von Übergangsformen kann nicht pauschal beantwortet werden. Dafür sind die Verhältnisse von Fall zu Fall zu verschieden. Markant unterschiedlich sind beispielsweise die Verhältnisse bei aktiv fliegenden Wirbeltieren. Während Fledermäuse von Beginn ihrer Fossilüberlieferung an unverkennbar ausgeprägte Fledermausmerkmale haben und sich stark von mutmasslichen Vorfahren unterscheiden (Rietbergen et al. 2023), ist die Situation bei den ersten Vögeln sehr viel anders. Überspitzt gesagt: Während der mutmassliche Übergangsbereich bei den Fledermäusen so gut wie leer ist, ist der Formenraum zwischen Dinosauriern und Vögeln so gut gefüllt, dass sich die Paläontologen – pointiert formuliert – vor Zwischenformen schier nicht retten können. Der berühmte «Urvogel» Archaeopteryx (Abb. 2) ist angesichts der heute bekannten Fülle von Fossilien, die Dinosaurier- und Vogelmerkmale kombinieren, nur ein Beispiel.

MEHRERE PROBLEME
Allerdings muss die Frage gestellt werden, ob alle diese Zwischenformen auch hinsichtlich ihrer Merkmale und der Reihenfolge ihres fossilen Auftretens wirklich als evolutionäre Übergangsformen in Frage kommen. Dabei zeichnen sich drei Probleme ab:
1. In Bezug auf einzelne Merkmale ist ein glatter Übergang zwischen Raubdinosauriern (Theropoden) und Vögeln nicht so deutlich, wie es evolutionstheoretisch zu erwarten wäre (Details bei Junker 2019). Dies gilt insbesondere für flugtaugliche echte Federn. Der Sprung von haarartigen, in manchen Fällen zerfaserten Körperbedeckungen zu flächigen Federn ist markant. Aber auch bei anderen Merkmalen finden sich solche Sprünge, wo sie aus funktionalen Gründen auch nicht verwunderlich sind (z. B. Fiederschwanz mit langer Schwanzwirbelsäule anstelle von Fächerschwanz mit Pygostyl = verwachsene Schwanzwirbel).
2. Die mutmasslichen Vorläufer- oder Übergangsformen passen in Bezug auf die zeitliche Reihenfolge ihres Auftretens sehr oft nicht in eine Evolutionsreihe (Junker 2019).
3. Die vogeltypischen Merkmale, die – wie behauptet – schrittweise evolutiv hinzugekommen sein sollen, treten nahezu durchweg mehrfach evolutionär unabhängig bzw. parallel auf. Evolutionstheoretisch muss somit angenommen werden, dass flächige Federn, das Pygostyl, der Hornschnabel und viele andere Merkmale mehrfach (bis siebenfach) unabhängig entstanden sind, weil sie bei verschiedenen «Stammbaumästen» unabhängig in verschiedenen Kombinationen mit anderen Merkmalen auftreten. Würde man die unterschiedlichen gemeinsamen Merkmale verbinden, entstünde ein dichtes Netzwerk statt eines Stammbaums (Beispiel: Abb. 3). Da aber fast jedes einzelne Merkmal der mutmasslichen Vogelevolution dabei einen alternativen Stammbaum ergeben würde, müssen sich Evolutionsbiologen für eine bestimmte Version von Abstammungsbeziehungen entscheiden und alle anderen Stammbaumrekonstruktionen verwerfen. Dies ist üblicherweise die Stammbaumversion, bei der die wenigsten unabhängigen Entstehungen baugleicher Merkmale (Konvergenzen) angenommen werden müssen.

Das bedeutet im Umkehrschluss, dass viele mögliche Abstammungsbeziehungen, die auf irgendwelchen gemeinsamen Merkmalen basieren, falsch wären. Die entsprechenden Zwischenformen wären damit keine Übergangsformen. Und aufgrund der ausgeprägten Vernetzungen (vgl. Abb. 3) und der häufigen Konvergenzen könnten sogar die meisten Zwischenformen nicht als Übergangsformen interpretiert werden. Das bedeutet aber nichts anderes, als dass es tatsächlich viel zu viele Zwischenformen, aber zu wenig passende Übergangsformen gibt.
Das zeigt beispielhaft: Man kann zwar immer Ähnlichkeitsbäume konstruieren, muss dabei aber in Kauf nehmen, dass Bäume sehr oft viele Widersprüchlichkeiten durch zahlreiche Konvergenzen aufweisen. Doch das ist evolutionstheoretisch problematisch, da bislang Konvergenzen evolutionär zu Recht als unwahrscheinlich angesehen wurden. Die mehrfach unabhängige Entstehung baugleicher Merkmale ist in einem zukunftsblinden Evolutionsprozess schliesslich nicht zu erwarten.
“Es gibt tatsächlich viel zu viele Zwischenformen, aber zu wenig passende Übergangsformen.
Der hier genannte Befund ist systematischer Art und nicht eine seltene Ausnahme: Die Ähnlichkeitsbeziehungen grösserer Gruppen von Tieren lassen sich aufgrund vieler Konvergenzen häufig besser durch ein Netzwerk darstellen als durch einen Baum (der als Stammbaum interpretiert werden könnte). Aus Platzgründen können im Folgenden dazu nur zwei weitere typische Beispiele grösserer Gruppen gebracht werden. 2
ZAHLREICHE WIDERSPRÜCHE
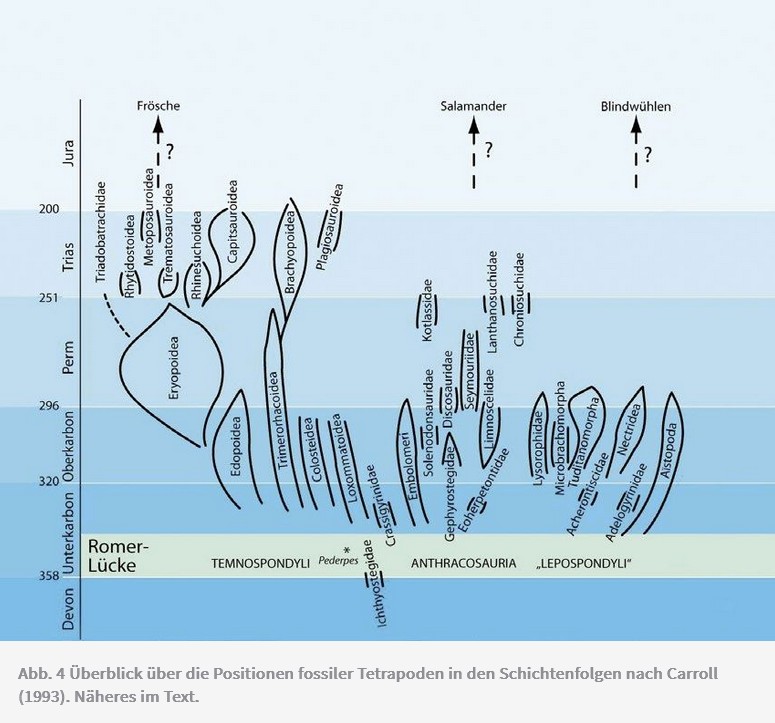
Bei den Amphibien- und Reptiliengruppen des Karbons (Abb. 4) gibt es ebenfalls zu viele Zwischenformen und zu wenig passende Übergangsformen für eine einigermassen widerspruchsfreie Baumdarstellung. Diese Formen treten mit unterschiedlichsten Merkmalskombinationen sogenannter «ursprünglicher» und «abgeleiteter» («höherentwickelter») Merkmale auf, sodass Verwandtschaftsverhältnisse laut Carroll (1992) kaum bestimmt werden können. Es müssen in grossem Ausmass Konvergenzen angenommen werden. Für keine Gruppe gibt es laut Carroll einen Konsens über die genauen Verwandtschaftsbeziehungen. Zudem gehören zu den ältesten karbonischen Fossilien ausgerechnet auch ausgesprochen abgeleitete Formen statt ursprüngliche Formen, wie man unter Annahme von Evolution erwarten würde. Fracasso (1994) benutzt das Bild von einer grossen «Wolke», die über den evolutionär anzunehmenden, aber nicht rekonstruierbaren Abstammungsbeziehungen hängt: «Eine Anzahl einzigartiger Linien ist zu erkennen, aber ihre Beziehungen bleiben verdeckt unter einer Wolke von verschiedenartigen und konvergenten Merkmalen ...» (Fracasso 1994, 118).
Beim zweiten Beispiel geht es um die Deuterostomier («Neumundtiere»). Bei diesen Tieren entwickelt sich in der Embryonalentwicklung der Urmund zum After und der definitive Mund entsteht auf der anderen Körperseite neu. Zu ihnen gehören vor allem die Stachelhäuter wie zum Beispiel Seesterne und Seeigel und die Chordatiere sowie einige kleinere Gruppen. Die zahlenmässig weitaus grösste Gruppe der Chordatiere sind die Wirbeltiere (Fische, Amphibien, Reptilien, Vögel und Säugetiere). Auch der Mensch gehört also zu den Deuterostomiern.
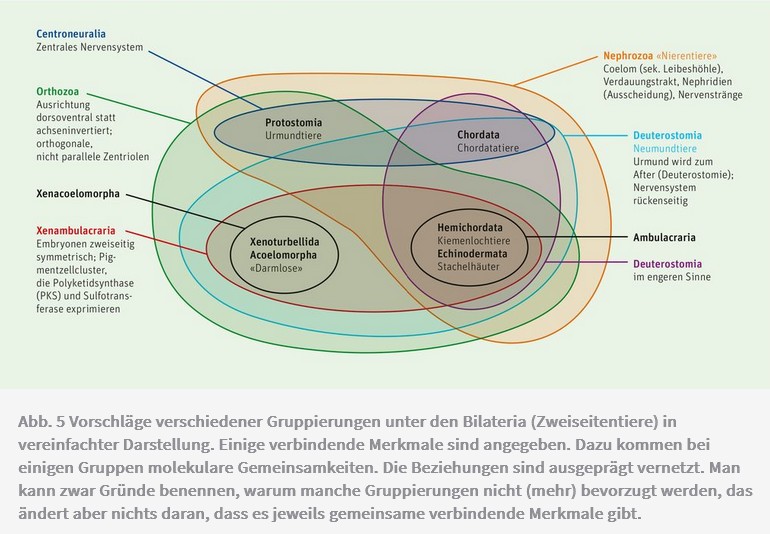
Eine Überblicksarbeit über den gegenwärtigen Stand des Wissens zum Ursprung der Deuterostomier von Nanglu und Kollegen (2023) stellt auch bei dieser hochrangigen Tiergruppe die Existenz vieler widersprüchlicher Merkmalskombinationen heraus. Als Konsequenz daraus wurden zahlreiche sich – je nach Merkmalsgewichtung – widersprechende Hypothesen über Verwandtschaftsverhältnisse aufgestellt, von denen keine als sicher belegt oder als klar widerlegt gelten kann (Abb. 5). Jede baumförmige bzw. eingeschachtelte Darstellung der Beziehungen der Deuterostomier-Gruppen ist dabei mit mehr oder weniger ausgeprägten Merkmalswidersprüchen behaftet. Zudem ist es schwierig, Merkmale zusammenzustellen, die allen Deuterostomiern gemeinsam sind. Die Autoren bezeichnen diese Situation bezüglich der Herkunft der Deuterostomier als «intellektuellen Wilden Westen». «Keine Hypothese, wie weit hergeholt sie auch erscheinen mag, kann völlig verworfen werden. Keine Theorie, und sei sie noch so verlockend logisch, kann von sich behaupten, sich gegen ihre Konkurrenten durchgesetzt zu haben» (Nanglu et al. 2023, 345).
Ein weiteres Problem ist: Die ältesten eindeutig zuordenbaren Fossilien der Tierstämme der Deuterostomier treten im Kambrium plötzlich als unterschiedliche Hauptlinien auf, die sich auch vom mutmasslichen letzten gemeinsamen Vorfahren der Deuterostomier beträchtlich unterscheiden (Nanglu et al. 2023, 318).
NETZWERK, KEIN STAMMBAUM
Als Muster von Merkmalsbeziehungen verschiedener Gruppen zeigt sich mit zunehmender Kenntnis von fossilen Arten mehr und mehr ein Netzwerk statt einem evolutionären Stammbaum. Das bedeutet: Je nach zugrunde gelegtem Merkmal sind verschiedene Arten oder höhere Gruppen einander am nächsten. Evolutionsbiologen geben dennoch ihre Stammbaumvorstellung nicht auf, müssen aber wohl oder übel akzeptieren, dass unerwarteterweise Konvergenzen und Merkmalswidersprüche sehr häufig ihren Stammbaumrekonstruktionen entgegenstehen.
Im Rahmen einer Schöpfungsanschauung kann man hingegen davon ausgehen, dass Merkmale frei kombinierbar sind und nicht bestimmten Evolutionspfaden folgen müssen. Netzartige Beziehungen von Gruppen von Lebewesen sind hier weder überraschend noch problematisch.
Reinhard Junker studierte Mathematik, Biologie und Theologie und promovierte in Interdisziplinärer Theologie. Nach kurzer Lehrertätigkeit war er 36 Jahre lang bis 2021 vollzeitlicher Mitarbeiter der «Studiengemeinschaft Wort und Wissen» und arbeitet dort weiterhin ehrenamtlich mit.
1 https://genesis-net.de/x/1-7/lueckenhafte-fossilueberlieferung/
2 In einem «Special Paper» habe ich eine grosse Anzahl ähnlicher Beispiele zusammengetragen (JUNKER 2025b).
www.factum-magazin.ch